Collaborators: Michael G. Frisk, Peter C. Wainwright, Bryan Kao,
John S. Sparks, F. James Rohlf

Skates are a diverse group of batoid fishes, comprising over 300 recognized species. A number of these species display a form of sexual dimorphism in which mature males a achieve what has been referred to as a ‘bell-shaped’ body. This is caused by a conspicuous concavity that develops in the anterior region of the pectoral fin as well as a generally more rounded posterior fin compared to females (Fig. 1). My work on this system has sought to identify a potential developmental mechanism driving pectoral fin dimorphism in skates and to assess its diversity within the group.
Based on previous work on cranial dimorphism in the bonnethead shark (Kajiura et al. 2005), I was interested in the ontogenetic development of pectoral dimorphism, particularly its relationship with sexual maturation. A study comparing sister species of skate in the genus Leucoraja, showed that males and females displayed divergent allometric trajectories with respect to pectoral shape and that much of the variation was due to morphological transformations in maturing males (Martinez et al. 2016). Pectoral fin dimorphism appeared to be due to elongation of male claspers, cartilage-supported intromittant organs. This was also observed in the bonnethead work and is proposed to have resulted from skeletal changes in the skeletal structures supporting male pectoral fins that are correlated with rapid elongation of clasper cartilages (Fig. 2). Interestingly, we do not see a similar pattern in rays (Myliobatiformes), presumably because their claspers are much smaller and less developed than that of skates (Fig. 3).


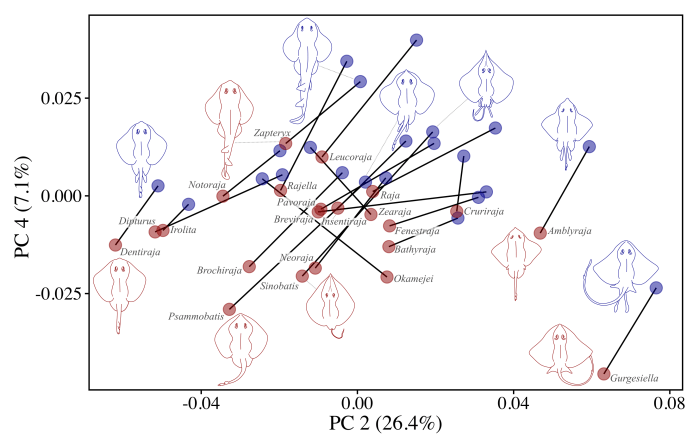
A more recent study across skate diversity, showed that pectoral dimorphism was a ubiquitous feature of the group (Martinez et al. in review), occurring in at least 131 species and 33 genera (e.g., Fig. 3) . In addition to skates, pectoral dimorphism was identified in a species of guitarfish, Zapteryx, suggesting a wider distribution outside of rajoids. While the nature of pectoral fin differences between males and females was largely consistent among species, the magnitude of dimorphic shape change varied by a factor of five across the group. Given the apparent link between pectoral dimorphism and maturation processes in skates, its expression may be indicative of the evolution of reproductive and life history strategies within the group.
Relevant Publications:
Martinez CM, Kao B, Sparks JS & Wainwright PC. 2019. Pectoral dimorphism is a pervasive feature of skate diversity and offers insight into their evolution. Integrative Organismal Biology. 1(1), 1-14.
Martinez CM, Rohlf FJ & Frisk MG. 2016. Sexual dimorphism in sister species of Leucorajas kate and its relationship to reproductive strategy and life history. Evolution & Development. 18(2), 105-115.
Funding Sources:
Lerner-Gray Fund for Marine Research (AMNH)
NSF Research Experience for Undergraduates, REU (AMNH; awarded to BK with mentors CMM and JSS)
